Nature Knows and Psionic Success
God provides
Anterior retrosplenial cortex is required for long-term object recognition memory
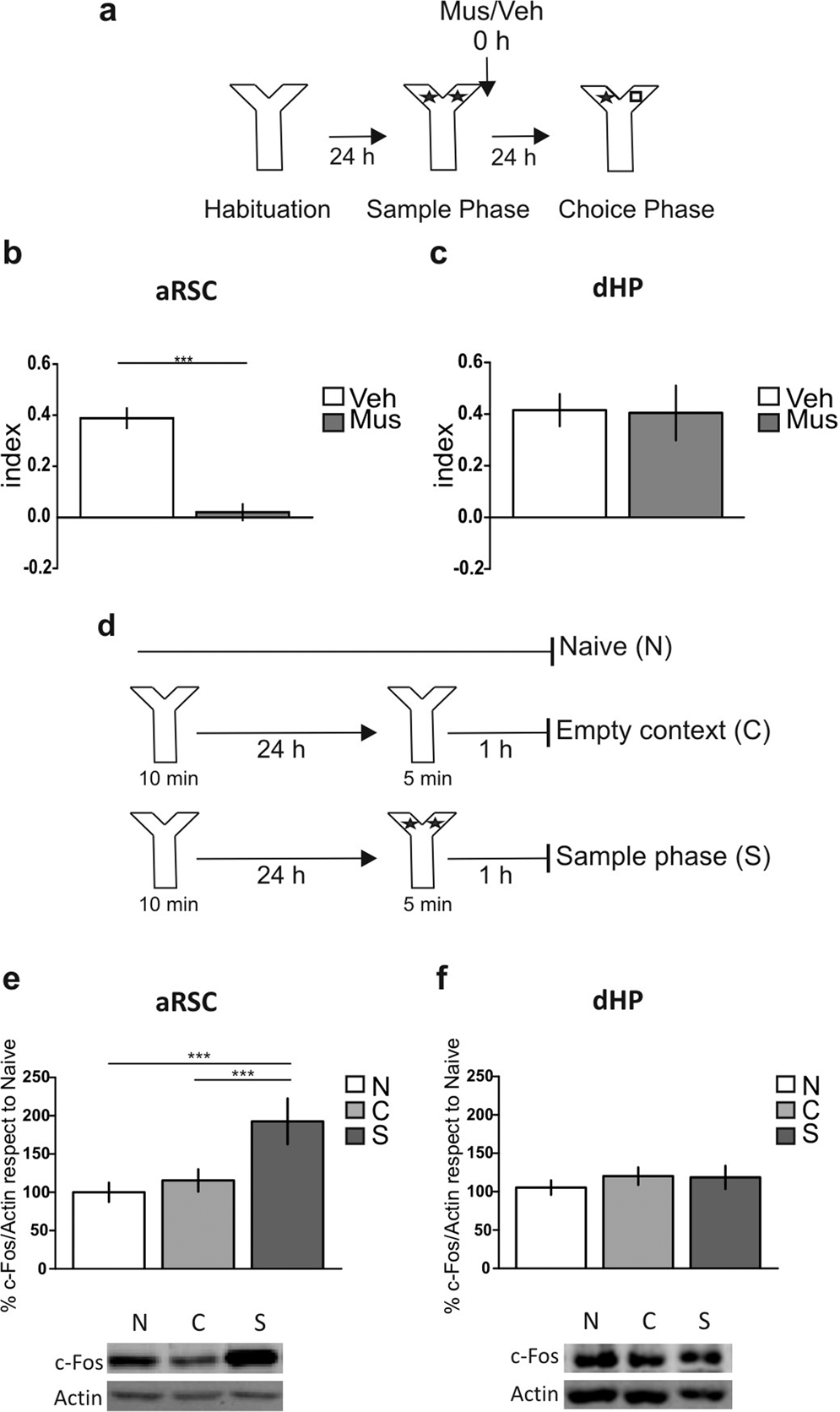
To make a comparative study about the participation of aRSC and dHP in the “what” component of the recognition memory, we decided to study object recognition memory in the non-spatial Y-maze (Y-OR) task. Due to the high walls, small corridors and the lack of explicit clues in the Y-maze, the Y-OR task has minimal spatial information and therefore a negligible “where” component. First, we validated the Y-OR task by transient inactivation of the PRh, confirming its requirement for object recognition memory (Supplementary Fig. ; p = 0.0031, t = 3.774, df = 11; Muscimol vs. Vehicle, Student’s t test, n = 5–7). Then, we analyzed the participation of the aRSC and dHP in the formation of Y-OR long-term memory (LTM) by infusing the GABA receptor agonist muscimol (0.1 μg/μl) or vehicle (saline) into the aRSC or the dHP immediately after the sample phase (training session), and we evaluated memory expression 24 h later during the choice phase (Fig. ). We found memory impairment associated to the aRSC inactivation (Fig. ; p < 0.0001, t = 6.611, df = 14; Muscimol vs. Vehicle, Student’s t test, n = 8 per group). In contrast, muscimol infusion into the dHP did not affect the Y-OR memory expression (Fig. ; p = 0.9278, t = 0.094, df = 7; Muscimol vs. Vehicle, Student’s t test, n = 4–5). To further evaluate the participation of these structures in Y-OR memory formation, we studied the levels of c-Fos expression, an immediately early gene that is a transcription factor usually upregulated in active brain structures, following a sample phase. Consistent with our behavioral data, c-Fos levels increased 1 h after the Y-OR sample phase in the aRSC (Fig. ; Bonferroni after two-way ANOVA, F = 0.31, p = 0.7347, F = 9.86, p = 0.0014, n = 4) but not in the dHP (Fig. ; two-way ANOVA, F = 0.27, p = 0.8977, F = 1.68, p = 0.2056, n = 3–5). Full-length blots are shown in the Supplementary Fig. .

The Power Within by Corey Daniels book available for only $2.99
Mind has both conscious and subconscious halves. These are likened to a driver and the truck he drives. The driver plans the destination and observes road conditions, while the truck provides motive power. Your subconscious mind is like the truck and it only goes in the direction in which is a steered. This can be the road or off a cliff. Likewise, the consciousness paints a picture of what the world is and what your goals are and the subconscious acts on them through emotion, physical response and energy, whether these are correct, rational images or false negative ones. The subconscious is also like an emotional reservoir which your body and mind draw responses from to external stimuli.
The Power Within lays out a method of programming your subconscious and tapping into the Holy Spirit, God voice or what the Greeks called the daimon (godman).